The transferable-signal gate: do learned policies beat their own init?
This is the first step of the larger inheritance-ladder experiment tracked in #904. That issue proposes building richer inherited payloads (P2 plasticity damping, P3 optimizer + replay-slice transfer, P4 gated/blended warm-start) on top of the existing P0 (Baldwinian) and P1 (Lamarckian) modes, then running a multi-seed A/B to find out when inheriting more of a parent’s learned state helps.
But none of that is worth building if there is nothing worth inheriting. The 05-16 “is the DQN actually learning?” post showed that within-life learning in the default config is barely detectable, and the 05-21 A/B and 06-04 newborn-level posts found no robust fitness gain from copying a parent’s policy. So #904 opens with an explicit precondition gate: in a learning-positive regime, does an end-of-life policy measurably make better decisions than a freshly-initialized one? If that budget is ~null, the richer-payload work stops and we record the negative result. This post runs that gate.
It passes — but only once the baseline is fair, and the real signal is much
smaller than it first looks. An early version of this gate compared the
trained policy against a pure-greedy random-init net and reported a huge
+566-616 reward budget. That number was an artifact: greedy-argmax on an
untrained network is a near-degenerate fixed-action controller that simply
dies, so the “budget” was almost entirely survival — “the trained net stays
alive, the broken one doesn’t.” Re-running against a non-degenerate baseline
(the real softmax(Q) x action_weights policy the agent actually uses) erases
the survival gap completely and leaves a genuine but modest decision-quality
signal: the trained policy forages a few percent better per step, worth ~+15-30
net reward over early life. That smaller signal is robust in all three profiles,
so the P2-P4 work is still justified — at a realistic effect size.
The regime
The gate deliberately does not use the default config, where within-life learning is ~null. It uses a learning-positive regime designed to give the DQN a fair chance to learn and to make the measurement clean:
- Small, fixed population (8 agents), long horizon (3000 steps). Long-lived agents accumulate a full lifetime of within-life learning, which is exactly the quantity the gate is about.
- Reproduction disabled. The runner patches
AgentCore.reproduceto a no-op while instrumentation is active. This isolates within-life learning from evolutionary dynamics (no inheritance, no selection) and keeps the run tractable — without it, high-resource ecologies have no population cap and the colony explodes into the hundreds, which both swamps the signal and exhausts RAM. - 6 seeds x 3 STABLE profiles. Seeds
{42, 7, 19, 101, 137, 256}acrossconservative/balanced/buffered, the same fixed-ecology profiles used by the inheritance A/B work, on thedevelopmentmap.
Runner: scripts/measure_transferable_signal.py.
Two-tier measurement
Measuring whether a policy changed is not the same as measuring whether it got better, so the gate reports both.
Tier 1 - probe-state drift (does the policy move at all?). Early in each run we cache a fixed set of observation tensors, snapshot every learning agent’s Q-net at birth and at end-of-life, and compare them on that frozen probe set: argmax disagreement, max-Q drift, and softmax KL. This is cheap and confirms the network actually moved, but movement alone is not improvement.
Tier 2 - grounded rollout differential (the decision metric). For each
harvested (birth, end-of-life) weight pair we run paired, single-agent
held-out rollouts and measure net RL reward as a differential
Δ = reward(end-of-life policy) − reward(init policy)
Two design choices make this measure decision quality rather than an artifact:
- A non-degenerate baseline. Both arms run the real action policy —
softmax(Q) x action_weightssampling — not pure-greedy argmax. This matters enormously (see below): under greedy, a random-init net collapses to one fixed action and dies, so “beats init” degenerates into “isn’t broken.” Under the weighted policy, a random-init net instead falls back to the chromosome action priors, i.e. the Baldwinian P0 baseline #904 actually compares against. - A survival-decoupled metric. Full-episode reward is
~survival_steps x foraging_rate, so it is dominated by how long the agent lives, not how well it decides. The gate therefore keys off early-age net reward (reward at a fixed age of 10/25/50 steps), plus per-step reward rate. Because every survivor accrues over the same short window, this isolates foraging/decision quality — exactly the “net early RL reward at ages N” the inheritance A/B is graded on.
Cohort aggregation uses the standard project robustness gate: a profile passes only if the paired (per-seed) 95% CI excludes zero and within-profile sign agreement >= 0.75.
Headline result: PASS (on the decision metric)
All three profiles clear both gate criteria on the survival-decoupled early-age reward metric, with perfect sign agreement:
| Profile | Mean Δ reward@age10 | 95% CI | Sign agr. | Robust | Mean drift |
|---|---|---|---|---|---|
| conservative | +15.6 | [12.3, 18.9] | 1.00 | yes | 0.80 |
| balanced | +17.8 | [14.2, 21.3] | 1.00 | yes | 0.87 |
| buffered | +18.1 | [15.5, 20.6] | 1.00 | yes | 0.83 |
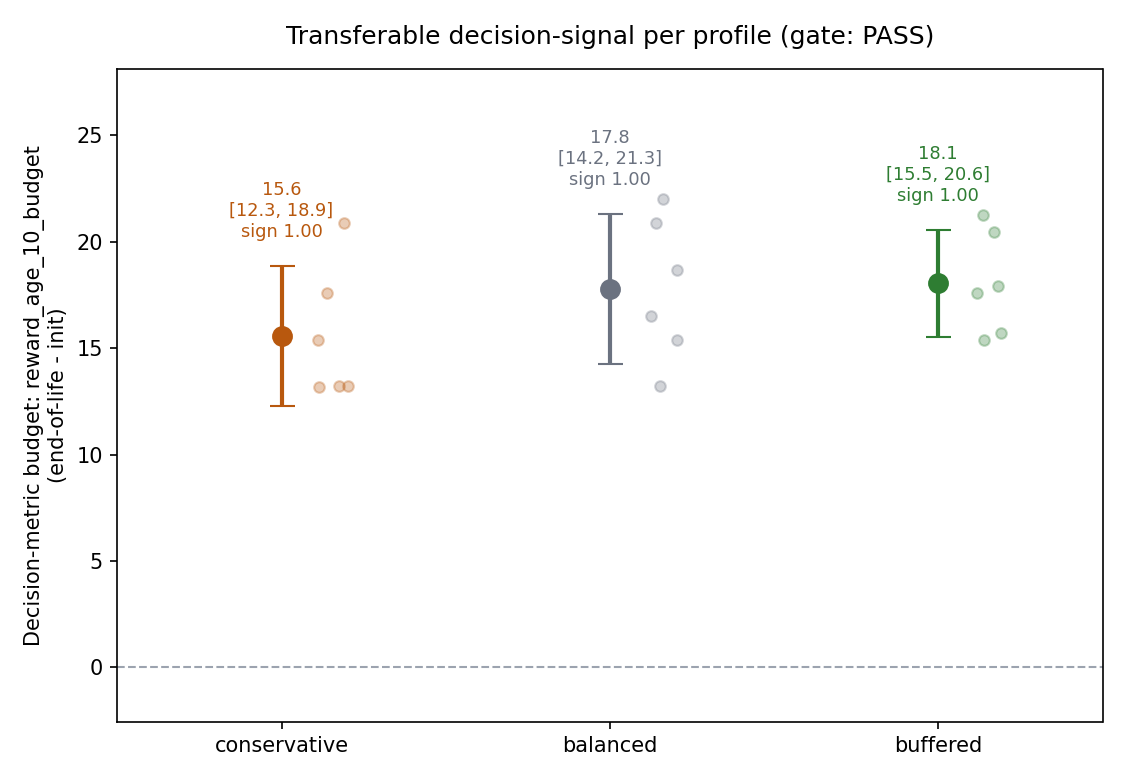
Every profile’s 95% CI sits above zero (faint per-seed points behind each marker), which is the verdict in one picture — note the y-axis tops out near 25, not 800.
Tier 1 agrees that the policies genuinely moved: mean argmax disagreement is 0.83 against a 0.05 threshold, with max-Q drift ~2.8-4.6 and softmax KL ~0.02-0.04 per cell. End-of-life policies are substantially different from their random init — and, on the fair baseline, that difference is a real (if small) improvement.
The baseline correction (why the number dropped 25x)
The first cut of this gate ran pure-greedy rollouts for both arms and reported a +566-616 budget that looked decisive. Two diagnostics killed it:
- Survival delta is now identically zero — for all 90 harvested policies. Under the realistic action-weighted policy, a random-init agent survives the held-out episode exactly as often as a trained one (both essentially always survive the easy single-agent ecology). The entire original budget lived in a survival gap that only existed because greedy-argmax on a random net is a degenerate controller that dies. Fix the baseline and the gap vanishes.
- Full-episode reward collapses from ~+587 to ~+24 (median +24.8, range -6.6 to +60.9; only 4/90 ≤ 0). What remains is not survival — survival is constant — it is a small genuine foraging edge.
So the lesson for the whole #904 program: the baseline, not the payload, was doing most of the talking. A win against a broken controller is not evidence that there is learned decision-quality signal to inherit.
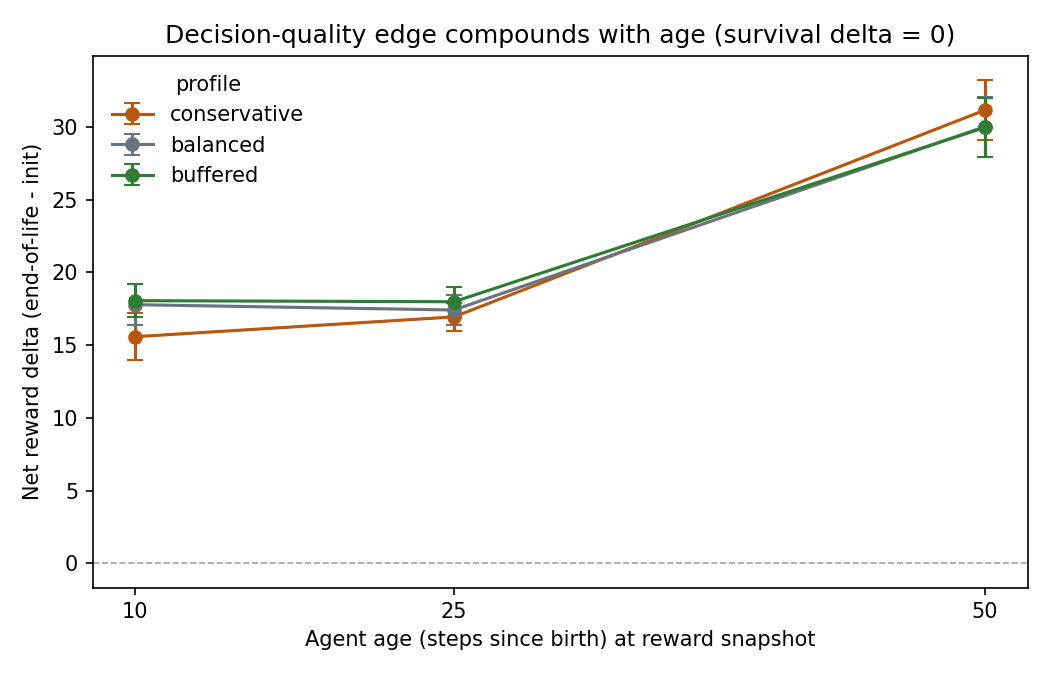
What the real signal looks like
With survival neutralized, the signal is a modest, consistent foraging improvement that compounds with age — the fingerprint of a genuinely better policy rather than noise:
- Net reward at age 10: +17.1 (99% of policies positive), age 25: +17.4 (100%), age 50: +30.4 (100%).
- Per-step reward rate +0.16/step (96% positive) — a few percent better than the ~5/step steady-state foraging rate.
The per-policy distribution of the gate metric is tight and almost entirely positive — no survival-driven ±800 bimodality, just a band a little above zero:
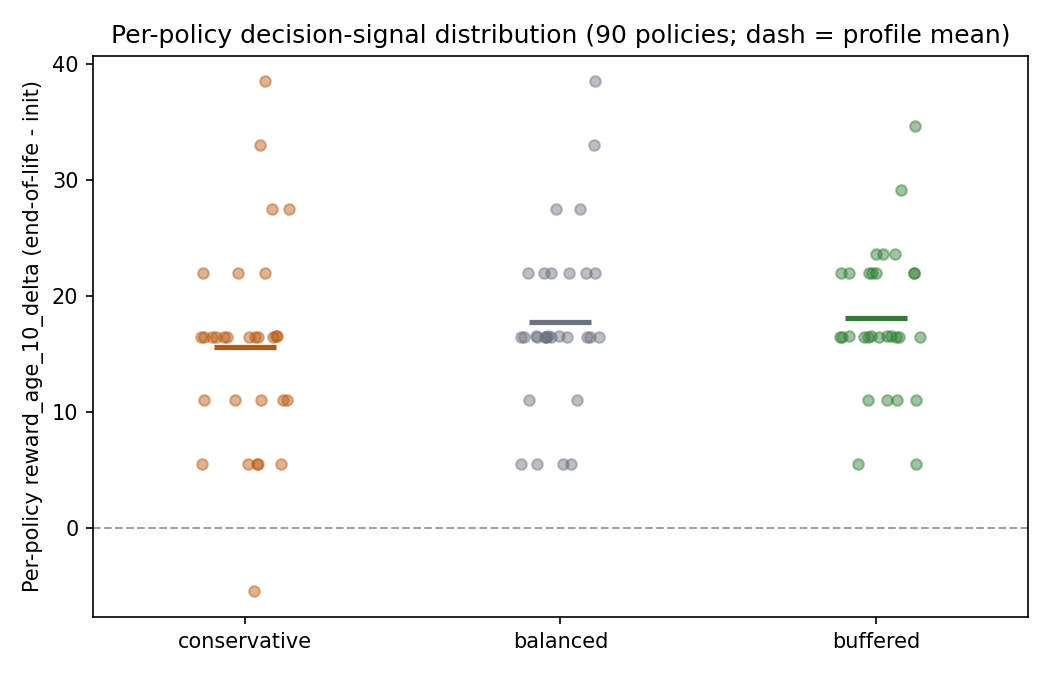
A few framing numbers:
- 90 harvested policies (5 per cell x 18 cells); harvested agents were long-lived (lifespans 1011-2999, mean ~2771 of 3000), so within-life learning had a full horizon to act.
- The signal is real but small: ~+0.16 reward/step is a single-digit-percent foraging gain, growing to ~+30 net reward by age 50. This is the honest size of the prize P2-P4 are competing for — not the +600 the broken baseline implied.
Patterns
- Across-profile means are tightly bunched at age 10 (15.6 / 17.8 / 18.1) and essentially converge by age 50 (~30 each), so the signal does not depend on a single favorable ecology.
bufferedis marginally strongest early (+18.1, narrowest CI) — more resources, slightly more reliable learning — but the gaps are within noise.- Seed 256 is the weakest (≈13 at age 10 in conservative/balanced) but still clearly positive, so it never threatens the gate.
What it means for #904
The gate passes on the right metric, so the precondition is satisfied and the P2-P4 inheritance ladder is worth building. But the corrected result reshapes expectations:
- Size the A/B for a small effect. The prize is ~+15-30 net early reward (~+0.16/step), not +600. The inheritance A/B should be powered to detect a single-digit-percent foraging gain and must report early-age net reward (the metric that survived scrutiny), never full-episode reward.
- Baselines matter more than payloads. The 25x drop here came entirely from making the baseline fair. P2-P4 must each beat the Baldwinian P0 policy (chromosome action priors), which is what a random-init net under the weighted policy already is — not a strawman. A richer payload that only beats a broken controller proves nothing.
- Don’t lean on survival. Under a fair baseline there is no survival
advantage to inherit in this regime; the transferable quantity is foraging
decision quality. Verify P2-P4 on that, and on cross-ecology retention (the
--cross-eval-profileprobe), not on “stays alive.”
What shipped + how to reproduce
The runner instruments a normal simulation rather than changing the core: it
monkeypatches the decision module to (a) capture the frozen probe set and each
agent’s birth/end-of-life Q-net snapshots during training, and (b) drive the
chosen eval policy during held-out evaluation. By default eval runs the real
action-weighted policy (--eval-policy weighted) and the gate keys off the
early-age reward budget (--gate-metric reward_age_10_budget); --eval-policy
greedy reproduces the old (degenerate) pure-greedy read for comparison. It also
pins torch to a single thread and blocks reproduction while active, so runs stay
fast and bounded.
The full 6x3 sweep (population 8, 3000 steps, eval horizon 150, 2 eval seeds,
reward ages 10/25/50, min_lifespan 100, top-5 policies per cell) reproduces
with:
PYTHONHASHSEED=0 python scripts/measure_transferable_signal.py \
--output-dir experiments/transferable_signal_v2
Outputs land in experiments/transferable_signal_v2/ —
signal_budget_summary.json / .md plus per-cell cell_result.json
checkpoints (re-run with --resume to skip completed cells). The three figures
above are regenerated with:
python scripts/plot_transferable_signal.py \
--summary experiments/transferable_signal_v2/signal_budget_summary.json
Open questions
- Does the small edge transfer without being washed out? The signal exists but is modest, which is exactly the regime where P1 (weights-only) was a null: a few good updates can be overwritten by a child’s first noisy steps. P2 (plasticity damping) and P3 (optimizer + replay) are the direct tests.
- Does it survive a cross-ecology shift? In-distribution the edge is robust;
the
--cross-eval-profileprobe asks whether a policy trained in one ecology still forages better in another, or whether it has overfit. That is the overfit discriminator for the payload ladder. - Which payload captures it? The gate says a (small) signal exists; #904’s ladder asks which of P2-P4 transfers it to offspring. That is the next post.